طراحی پرایمر و پروب اختصاصی
پرایمر
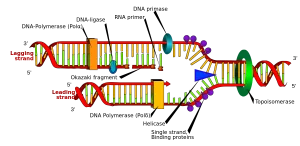
پرایمر رشته کوتاهی از RNA یا DNA است که معمولاً از ۱۸ تا ۲۲ باز تشکیل شده است. این رشته کوتاه به عنوان نقطه ای برای آغاز سنتز DNA عمل می کند. پرایمرها معمولاً الیگونوکلئوتیدهایی کوتاه هستند که به روش شیمیایی سنتز شده اند. آن ها به DNAی هدف متصل می شوند که پس از آن فرآیند نسخه برداری توسط پلیمراز قابل انجام می شود. پرایمر جزئی ضروری در پروسه رونویسی DNA به شمار می رود، زیرا آنزیمهای کاتالیز کننده این فرآیند (DNA polymerase ها) تنها می توانند نوکلئوتیدها را به یک رشته DNA اضافه کنند. آنزیم های پلیمراز، رونویسی را از پایان ‘ 3 پرایمر شروع کرده و به این روش به کپی کردن رشته مقابل DNA می پردازد. در رونویسی DNA در شرایط طبیعی، از رشتههایی از جنس RNA که به آن ها پرایمر RNA گفته می شود، به منظور سنتز DNA در هر دو رشته پیشرو و پسرو استفاده می گردد.
از سوی دیگر بسیاری از روش های آزمایشگاهی که نیازمند آنزیم DNA پلیمراز هستند، از پرایمرهای DNA استفاده می کنند زیرا که پرایمرهای DNA مقاومت دمایی بیشتری دارند.
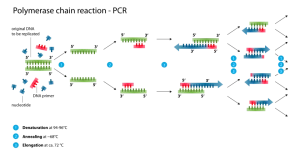
از این تکنیکها میتوان به توالی یابی دی ان ای (DNA sequencing) و واکنش زنجیره ای پلیمراز (polymerase chain reaction) اشاره کرد.
در آزمایشات، پرایمری که دمای ذوبش به دمای اتصال آن نزدیک باشد، اهمیت بسیار دارد زیرا امکان اتصال به محل نادرست در یک پرایمر که دمای ذوبش به طور معنی دار بالاتر از دمای واکنش اتصال (annealing) است، وجود دارد که این امر منجر به اشتباه در رونویسی شدن توالی می شود. این در حالی است که اگر دمای ذوب پرایمر به طور معنی دار پایین تر از دمای اتصال رشته باشد، احتمالا نتواند به مکان مورد نظر بچسبد و قطعه مورد نطر تکثیر نمی شود.
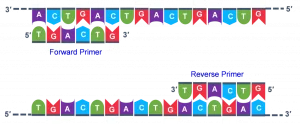
طراحی پرایمر صحیح برای درست انجام شدن یک واکنش PCR بسیار ضروری است. برای تکثیر یک قطعه با بهره وری بالا رعایت نکاتی ضروری است:
- طول پرایمر
به طور معمول، طول ۲۲-۱۸ نوکلئوتید برای یک پرایمر بهینه است. اگر طول پرایمر کمتر باشد، احتمال اتصال غیر اختصاصی بالا می رود و اگر از این محدوده بیشتر باشد، اتصال پرایمر به الگو در دمای اتصال سخت تر می شود.
- دمای ذوب پرایمرها
دمای ذوب پرایمرها یا به اصطلاح Tm، دمایی است که نیمی از DNA دو رشته ای از هم جدا می شود و DNA تک رشته ای ایجاد می کند. Tm نشانگر پایداری DNA دورشته ای است. پرایمرهایی با Tm 58-52 درجه ی سانتی گراد معمولا بهترین نتایج را به دست می دهند. Tm متأثر از محتوای GC یک توالی است.
- دمای اتصال پرایمرها
دمای ذوب پرایمرها تخمینی از میزان پایداری هیبرید DNA-DNA است و در تعیین دمای اتصال خیلی اهمیت دارد. اگر دمای اتصال(Ta) خیلی بالا باشد، پرایمرها و الگو از هم جدا باقی می مانند و اتصال صورت نمی گیرد و اگر دمای اتصال خیلی پایین باشد تنها برخی از نوکلئوتیدهای پرایمرها به الگو متصل می شود و اتصال درستی رخ نمی دهد و محصولات غیر اختصاصی تولید می شود. از فرمول Rychlik برای به دست آوردن Ta استفاده می شود:
Ta= 0.3×Tm(primer)+ 0.7×Tm(product)−14.9
- محتوایGC
نسبت بازهای GC به کل بازها باید بین ۶۰-۴۰ درصد باشد.
- گیره GC (GC clamp)
وجود بازهای GC در ۵ باز انتهای ﹱ۳ پرایمرها (که به آن گیره GC گفته می شود) اتصال اختصاصی در انتهای ﹱ۳ را بهبود می بخشد (به علت اتصال قوی تر بازهای G و C. در ۵ باز انتهایی سمت ﹱ۳ نباید بیش از 3 G یا C قرار داشته باشد.
- ساختار ثانویه پرایمر
حضور ساختارهای ثانویه ای که تحت تاثیر برهم کنش های بین مولکولی یا درون مولکولی ایجاد می شوند، منجر به تولید محصول بسیار ناچیز PCR می شود و یا اصلا محصولی تولید نمی شود. وجود این ساختارهای ثانویه برخلاف اتصال پرایمرها به الگو و تکثیر عمل می کنند. این ساختارها به میزان قابل توجهی دسترسی به پرایمرها را کاهش می دهند.
- ساختارهای سنجاق سری(hairpins)
این ساختارها جزء برهم کنش های درون مولکولی پرایمرها هستند و باید از آن ها دوری شود. اگر انتهای ﹱ۳ پرایمر یک ساختار سنجاق سری با ΔG=-2 Kcal/mol داشته باشد (انرژی آزاد گیبس، هرچه منفی تر باشد، ساختار پایدارتر است) و یا یک ساختار سنجاق سری درونی با ΔG= -3 kcal/mol داشته باشد، معمولا قابل تحمل است و مشکلی ایجاد نمی کند.
- دیمر خودی(self dimer)
دیمر خودی پرایمر ناشی از برهم کنش های بین مولکول های پرایمر (از یک جنس) است. معمولا در یک واکنش از مقادیر زیادی از پرایمرها نسبت به میزان الگوی PCR استفاده می شود.
وقتی پرایمرها تمایل شان برای اتصال به خود بیشتر از تمایل شان برای اتصال به الگو باشد، محصول PCR کمتری تولید می شود.
معمولا دیمر انتهای ﹱ۳ با ΔG= -5 kcal/mol و دیمرهای درونی با ΔG= -6 kcal/mol قابل تحمل هستند.
- دیمر متقابل(cross dimer)
دیمرهایی که براساس برهم کنش های بین مولکولی بین پرایمرهای سنس و آنتی سنس (جلویی و عقبی) به وجود می آیند.
به طور معمول دیمرهای متقابل انتهای ﹱ۳ با ΔG= -5 kcal/mol و دیمرهای متقابل درونی با ΔG= -6 kcal/mol قابل تحمل هستند.
- تکرارها
شامل تکرارهای دو نوکلئوتیدی که مدام در یک توالی تکرار می شوند. حداکثر تعداد تکرارهای قابل قبول، ۴ تکرار دو نوکلئوتیدی است.
- Runs
منظور پرایمرهایی با تکرارهای تک نوکلئوتیدی زیاد، پشت سر هم است که می تواند باعث اتصال غیر اختصاصی پرایمر شود. به عنوان مثال AGCGGGGGATGGGG تکرارهای ۵ و ۴ تایی از G را دارد. حداکثر تعداد تکرار قابل قبول برای یک باز، ۴ جفت باز است.
- پایداری انتهای ﹱ۳
منظور حداکثر میزان ΔG برای ۵ باز انتهایی ﹱ۳ است. انتهای ﹱ۳ نا پایدار (با ΔG منفی کمتر) منجر به اتصال های اشتباه کمتری می شود.
از ساختارهای ثانویه الگو دوری کنید:
پرایمرها باید برای ناحیه ای طراحی شوند که آن ناحیه، ساختار ثانویه پایداری تشکیل ندهد. در غیر این صورت این ساختارهای ثانویه پایدار اجازه اتصال پرایمرها به الگو را نمی دهند.
- از همولوژی متقابل دوری کنید
برای بالا بردن اختصاصیت پرایمرها باید از نواحی همولوگ دوری کرد. به عبارت ساده تر، پرایمرها باید به گونه ای طراحی شوند که نواحی دیگر ژنومی را نشناسند و آن ها را تکثیر نکنند. با BLAST کردن پرایمرها این ویژگی بررسی می شود.
- طول قطعه تکثیر شونده
طول قطعه تکثیر شونده در qPCR نزدیک به ۱۰۰جفت باز و در PCR استاندارد نزدیک به ۵۰۰ جفت باز است. اگر جایگاه قرارگیری پرایمرها بر روی الگو را بدانید، طول محصول از فرمول زیر به دست می آید:
- طول محصول= (جایگاه پرایمر جلویی- جایگاه پرایمر عقبی)+۱
- دمای ذوب جفت پرایمرها
یک جفت پرایمر(عقبی و جلویی) باید Tm نزدیک به هم داشته باشند تا بهترین کارآیی را داشته باشند. اختلاف ۵ درجه سانتی گراد یا بیشتر مانع تکثیر می شود.
انواع پرایمرهای پرکاربرد
1- In silico Primers: ارزیابی پرایمرها قبل از سنتز آنها
2- Standard Primers: تکثیر آمپلی کون مورد نظر بدون هیچ حذف و اضافه ای ،تشخیص ویروس و باکتری
3- Degenerate Primers: شناسایی توالی نوکلئوتیدی ساختارهای کدکننده پروتئین ها و پپتیدها
4- Inverse Primers: شناسایی موقعیت insert ژنومی یا سکانس شناخته شده ای در ژنوم ،مطالعه متاژنوم
5- cDNA Primers or RT-Specific Primers : شناسایی ویروس های RNAدار ،تعیین مقدار ویروس موجود در بدن موجود زنده ، بررسی سطح بیان ژنها
6- Cloning Specific primers : آماده سازی insertهابرای وارد نمودن در وکتور مناسب
7- Linker Primers : تکثیر توالی های شناسایی نشده ، آماده سازی توالی های شناسایی نشده برای توالی یابی
8- Assembly Primers : سنتز توالی های طویل DNA ، طراحی و تولید بیوسنسورها
9- Asymmetric Primers : توالی یابی ، ساخت پروب ، آپتامر
10- Multiplex Primers : تکثیر و شناسایی توالی های متعدد با انجام یک واکنش
11- MLPA Primers : تکثیر و شناسایی توالی های متعدد با یک جفت پرایمر با انجام یک یا چند واکنش
12- Nested Primers : تکثیر اختصاصی تر آمپلی کون نسبت به نوع استاندارد ، کاهش زمینه نامطلوب تکثیر غیراختصاصی DNA ، افزایش مقادیر کم الگو
13- SOEing Primers : اتصال اگزانها به هم و خارج سازی اینترانها ، ایجاد ساختارهای کایمریک
14- پرایمرهای ARMS: شناسایی جهش یا بیماری ژنتیکی ناشی از تغییر توالی نوکلئوتیدی ، تعیین نوع SNP
15- پرایمرهای Dial-out : بازیابی مولکولهای DNA از کتابخانه DNA
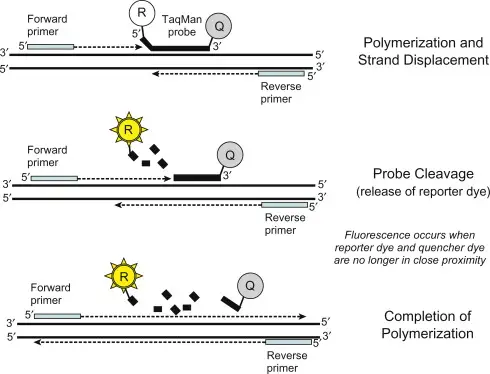
پروب
در انجام تحقیقات ژنتیکی، اغلب از پروب استفاده می شود. پروب ها(کاوشگر) امتدادی از DNA یا RNA هستند که یک برچسب به آنها چسبانده می شود. این برچسب به ما اجازه می دهد تا ببینیم DNA در یک سلول، یا در کروموزوم یا حتی در DNA خالص جدا شده کجا متصل میشود.
پروب ها را با مولکول های مختلف می توان برچسب گذاری کرد. می توان از مواد رادیواکتیو یا مواد فلورسنت برای اتصال شیمیایی آن به یک کاوشگر استفاده کرد.سپس می توان از آن کاوشگر برای جستجوی جایی که mRNA های خاصی در یک سلول یا در یک بافت بیان می شوند استفاده کرد.
همچنین می توان از کاوشگرها برای غربالگری ژنوم استفاده کرد تا بفهمیم آیا نسخه های اضافی وجود دارد، که اغلب در سرطان ها اتفاق می افتد، یا نسخه های از دست رفته بخش های خاصی از ژنوم وجود دارد، که در سندروم های ارثی و در سرطان ها اتفاق می افتد.
منابع
- Binas M (2000). Designing PCR primers on the web. Biotechniques 29: 988-990.[Pubmed]
- Dieffenbach CW, Lowe TMJ, Dveksler GS (1995). General Concepts for PCR Primer Design. In: PCR Primer, A Laboratory Manual, Dieffenbach CW, Dveksler GS Ed., Cold Spring Harbor Laboratory Press, New York, 133-155.
- Erlich HA, Gelfand D, Sninsky JJ (1991). Recent advances in the polymerase chain reaction. Science 252: 1643-1651. [Pubmed]
- Lowe T, Sharefkin J, Yang SQ, Dieffenbach CW (1990). A computer program for selection of oligonucleotide primers for polymerase chain reaction. Nucleic Acids Res. 18: 1757-1761. [Pubmed]
- با سپاس و قدردانی فراوان از مجموعه زیست ژن بافت




